|
 Australoporus tasmanicus Australoporus tasmanicus
SynonymsFlaviporus aroha
Heterobasidion tasmanicum
Poria aroha
Fomes cuneatus
Fomitopsis cuneata
Fomitopsis tasmanica
Trichaptum tasmanicum
Polyporus tasmanicus
BiostatusPresent in region - Indigenous. Non endemic
Images (click to enlarge)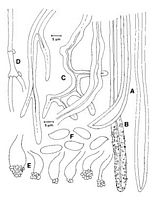
Caption: Fig. 1. Australoporus tasmanicus. A,C, skeletal hyphae from the ontext; B, encrusted skeletal hypha
from the dissepiments; D generative hyphae; E, ventricose cystidia from the hymenium; F, spores.
From the type of Poria aroha (PDD 5254). | 
Caption: specimen RR NZ04
Owner: Richard Robinson | 
Caption: Australoporus tasmanicus
Owner: Nils Hallenberg | 
Caption: Australoporus tasmanicus
Owner: Nils Hallenberg | 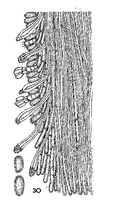
Caption: Flaviporus aroha x 600, spores x 1200. Section through part of a dissepiment showing skeletal and generative hyphae, the latter without clamp connections, metuloids with apices crystal encrusted, and spores.
Owner: Cunningham, G.H. |  |
Article: Buchanan, P.K.; Ryvarden, L. (1988). Type studies in the Polyporaceae - 18. Species described by G.H. Cunningham. Mycotaxon 31(1): 1-38.
Notes: Cunningham (1965) transferred the species to Flaviporus Murr., while stating that apart from the
presence of 'metuloids' the species could belong in Heterobasidion Bref. sensu Cunn. Comparison
of the type specimens of Poria aroha, Polyporus tasmanicus Berk. (K), Fomes cuneatus Lloyd (K),
and Polyporus suaderis Lloyd (K) show that all are conspecific, with P. tasmanicus being the oldest
name.
Macroscopically, the species varies from resupinate to distinctly pileate. The perennial, narrowly
concentrically sulcate and zonate, often ungulate pileus and the pale pink to orange pore surface,
when fresh, are diagnostic characters.
In the descriptions of both H. tasmanicum and Flaviporus aroha, Cunningham (1965) mistakenly
reported generative hyphae to be simple septate when, in fact, they have clamps. He also omitted to
report that skeletal hyphae are strongly dextrinoid, becoming red-brown in Melzer's reagent.
Skeletal hyphae often protrude into the hymenium, and at pore mouths the ends of skeletal hyphae
are finely encrusted. The thick-walled metuloids described for F. aroha were found to be thin-walled, sometimes collapsed, apically encrusted cystidioles. They are more abundant in fertile
specimens and may be scarce or absent in old specimens with a poorly preserved hymenium. Spores
are elongate ellipsoid, smooth- and thin-walled, nonamyloid, nondextrinoid, and hyaline in the
tubes. Those observed on the pileus surface however are light brown and often larger than those in
the tubes. Average spore measurements are 8.5-13 x 3.5-6.5 µm.
The unusual combination of characters found in P. tasmanicus means that the species cannot be
accommodated in any of the four genera to which Cunningham assigned it. Indeed, we have found
no suitable genus for this species.
Dextrinoid skeletal hyphae are found in species of Perenniporia Murr., but this genus has distinctly
truncate, thick-walled spores with a variable dextrinoid reaction and lacks encrusted skeletal
hyphae. Navisporus Ryv. is a close relative, having dextrinoid skeletal hyphae and navicular to
cylindrical spores of similar size to those in Polyporus tasmanicus. However, skeletal hyphae are
smooth and no encrusted cystidioles are known in the two species so far described in the genus.
Furthermore, the species are brown in colour and have a loose consistency, very different from the
hard, light-coloured fruit-bodies of P. tasmanicus. Junghuhnia Corda is characterised by a dimitic
hyphal system of a similar type to that found in P. tasmanicus. Cystidia are present, but are
developed from the skeletal hyphae and are encrusted at the apex. Thin-walled, ventricose cystidia
have not been observed in this genus and the skeletal hyphae are not dextrinoid.
Since this species cannot be accommodated in any known genus without changing the concepts of
that genus, we propose a new genus for P. tasmanicus [Australoporus].
Article: Gadgil, P.D. (in association with Dick, M.A.; Hood, I.A.; Pennycook, S.R.) (2005). Fungi on trees and shrubs in New Zealand. Fungi of New Zealand. Ngā Harore o Aotearoa 4: xi + 437 p. Hong Kong: Fungal Diversity Press.
Description: Type: Lignicolous Fungi; Description: Basidiomata perennial, solitary or with pilei overlapping each other, firm and woody, attached by a broad lateral base. Pileus effused-reflexed, sometimes ungulate, applanate, or resupinate, 70–170 mm wide, 30–50 mm thick. Pileus surface concentrically sulcate and ridged, radially fluted, light orange brown, darkening to umber with age. Pore surface with a sterile border 2–3 mm wide, white, becoming dirty brown, pores obscurely stratose, 5–6 per mm. Context pale fawn to ochre, 2–5 mm thick. Hyphal system dimitic. Basidiospores elliptical, 7–10 × 4–5 μm, smooth, hyaline.
Distribution: Distribution: Northland, Auckland, Coromandel, Waikato, Bay of Plenty, Taupo, Wellington, Nelson, Buller, Westland, Fiordland, North Canterbury, Otago Lakes, Southland.; 1st Record: Lloyd (1915: as Fomes cuneatus).
Article: Cunningham, G.H. (1965). Polyporaceae of New Zealand. New Zealand Department of Scientific and Industrial Research, Bulletin 164: 304 p. Wellington:.
Description: Hymenophore perennial, solitary, coriaceous, seldom pileate, commonly resupinate and loosely attached. Pilei merely upturned edges of one margin of broadly resupinate areas, with a radius of 8 mm; when resupinate effused to 16 x 5 cm, commonly 2-4 mm thick (in one specimen to 25 mm), with several outlying islands; pileus surface deep chestnut or umber, finely velutinate, concentrically sulcate; cortex to 200 µm thick, ferruginous, of mainly vertical hyphae compacted and partly cemented and with walls ferruginous; margin (of pilei) bluntly acute, concolorous, crenate (of resupinate specimens), irregular, thinning out, finely tomentose, lifting; hymenial surface white, drying cream or pallid tan, even, or undulating and nodose, each layer receding. Pores in 2-6 layers with context hyphae between, commonly in one or two layers, pallid isabelline, to 4 mm deep in each layer, round or angular, 4-5 per mm, 100-150 µm diameter; dissepiments 50-150 µm thick, occasionally to 500 µm, apices tapering, even, finely velutinate with erect hyphae, ends of which are crystal encrusted. Context 1-2 mm thick, straw or tan, closely intertwined and with coloured mucilage between hyphae; skeletal hyphae to 4 µm diameter, lumena 1-2 µm wide, aseptate, sparsely branched, irregularly waved towards ends, walls staining; generative hyphae to 2.5 µm diameter, walls 0.2 µm thick, somewhat sparingly branched and septate. Hymenial layer to 25 µm deep, a scanty palisade of basidia, paraphyses, paraphysate hyphae, and metuloids. Paraphysate hyphae merely projecting ends of skeletal hyphae, with thin walls encrusted with fine crystals. Metuloids clavate or fusiform, to 25 x 10 µm, walls hyaline or tinted, to 3 µm thick, capped with coarse crystals. Basidia clavate, 12-16 x 3.5-4 µm, bearing 4 spores; sterigmata erect, to 5 µm long. Paraphyses subclavate, 8-12 x 3-3.5 µm. Spores elliptical, apiculate, 9-12 x 4.5-6 µm, walls smooth, hyaline, 0.1 µm thick, soon collapsing.
Habitat: Bark or decorticated wood of dead fallen trunks, associated with a white rot.
Distribution: New Zealand.
Notes: Most collections are resupinate with one or several receding layers. In two, pilei are present, appearing as scantily developed margins of upper edges of plants growing vertically upon fallen logs. A well developed cortex is present in them, indicating that they are genuine pilei. When fresh the hymenial surface is white, although in some specimens it may be isabelline, changing in drying to some shade of brown. One collection taken from the under side of a fallen log was 1.5 m long, 2.5 cm thick, and composed of six receding layers. When fresh, plants possess a fragrant odour as of ripe peaches. Metuloids are scanty, but readily seen as they project to 10 µm, possess thick and refractive, usually tinted walls, and are capped with coarse crystals. Paraphysate hyphae are also present, both in the hymenial layer and at apices of dissepiments. The species is somewhat difficult to place. Other than for the presence of metuloids, it is a typical Heterobasidion; the presence of metuloids places it under Flaviporus with which it agrees in several features.
Article: Cunningham, G.H. (1965). Polyporaceae of New Zealand. New Zealand Department of Scientific and Industrial Research, Bulletin 164: 304 p. Wellington:.
Description: Comb. nov.
Hymenophore perennial, solitary or imbricate, firm and woody, attached by a broad lateral base. Pilei usually effused-reflexed, sometimes ungulate or applanate, occasionally resupinate, 7-17 cm, 2-5 cm radius, 3-5 cm thick; pileus surface bay, becoming umber, concentrically sulcate and ridged, radiately fluted and/or rugulose, nodose; cortex to 0.5 mm thick, dark brown, shining, of compactly intertwined cemented hyphae; margin acute, entire, undulate, lighter in colour; hymenial surface ochre drying bay, with a sterile border 2-3 mm wide. Pores obscurely stratose, pallid ochre, 2-4 mm deep in each layer, 5-6 per mm, 100-200 µ diameter, dissepiments 50-150 µ thick, most 50-75 µ, apices expanded partly occluding pores, velutinate. Context 2-5 mm thick, ochre or isabelline, of compact intertwined hyphae; skeletal hyphae to 4 µ diameter, lumena almost capillary, sparsely branched, aseptate, staining; generative hyphae to 3 µ diameter, walls 0.2 µ thick, branched, septate. Hymenial layer to 20 µ deep, a close palisade of basidia and paraphyses soon collapsing and partly embedded in mucilage. Basidia clavate, 12-16 x 7-8 µ, bearing 4 spores; sterigmata erect, to 4 µ long. Paraphyses clavate, 8-12 x 5-6 µ. Spores elliptical, 7-10 x 4-5.5 µ, walls smooth, hyaline, 0.1 µ thick, staining slightly.
Habitat: Bark or decorticated wood of dead erect trunks, associated with a white rot.
Distribution: Australia, Tasmania, New Zealand.
Notes: Plants sometimes exude large drops from the hymenial surface, and occasionally these persist so that pore tissue grows between them; consequently when drops disappear eventually the surface exhibits a pitted or reticulated appearance. From Heterobasidion hemitephrum the species may be separated by the different shape and surface of the pileus, larger spores and absence of an orange zone beneath the cortex. Spores vary in length in different specimens, some not exceeding 6.8 x 3.5-4 µ, in others reaching a length of 10 µ.
Article: Cunningham, G.H. (1947). New Zealand Polyporaceae. 1. The genus Poria. New Zealand Department of Scientific and Industrial Research, Plant Diseases Division, Bulletin 72: 43 p.
Description: Hymenophore effused, to 16 x 5 cm., 2-4 mm. thick, firm with numerous outlying islands;
surface cream or isabelline, or occasionally drying some shade of brown, even, or undulating and
nodose ; margin irregular, rounded, waved, finely tomentose, byssoid, sterile for a width of 2-3
mm., lifting. Pores not in strata, round or angular, often oblique, in section pallid isabelline, to 4
mm. deep, 100-150 µ diameter, or 4-5 to mm.; dissepiments 50-150 µ, occasionally to 500 µ
thick, equal, of woven partly parallel hyphae, apices tapering, firmly woven, a few apical hyphae
with rounded apices and crystal coated. Context 1-2 mm. thick, floccose, pallid isabelline,
closely woven, not gelatinized but with coloured gelatinous matter scattered between hyphae;
skeletal hyphae to 4 µ thick, lumen 1-2 µ, staining less deeply in old hyphae which are more
thickened in walls, aseptate, sparsely branched, somewhat irregularly waved near ends;
generative hyphae to 2 µ thick, delicate walled, tinted by stains, somewhat sparingly branched
and septate, readily collapsing, clamp connections present but scanty. Cystidia present in the
hymenium, hyaline or tinted yellow, clavate, to 25 x 10 µ, walls about 3 µ thick, capped with
crystals. Basidial type clavate, basidia clavate, scanty, to 16 x 1 µ, soon collapsing. Spores
elliptical, 9-12 x 4-6 µ, hyaline, smooth, soon collapsing.
Distribution: New Zealand.
Notes: When fresh plants are white or cream, though a few may be coloured isabelline. Spores are
present only in young plants, since basidia and spores early collapse, leaving only remnants
attached to walls of dissepiments. Specimens possess a pleasant smell when freshly gathered, as
of ripe peaches; this soon disappears when plants are dried. The large smooth spores separate the
species from all save P. leucoplaca. From this it differs in the presence of capitate cystidia;
thicker differently coloured hymenophore and scantily branched skeletal hyphae.
Article: Cunningham, G.H. (1948). New Zealand Polyporaceae. 5. The genus Fomitopsis. New Zealand Department of Scientific and Industrial Research, Plant Diseases Division, Bulletin 76: 8 p.
Description: Hymenophore perennial, dimidiate, sometimes imbricate, firm and woody. Pileus effused-reflexed, applanate or ungulate, sometimes resupinate, 7-17 cm. x 2-5 cm. x 3-5 cm.; surface concentrically sulcate and ridged, fluted, nodose, bay brown, becoming umber, cuticle to 0.5 mm. thick, dark brown, shining, of compacted mainly woven hyphae cemented together; margin acute, entire, undulate, lighter in colour; hymenial surface at first ochraceous, drying bay brown, concave, sterile margin 2-3 mm. wide, finely tomentose, dissepiments not toothed. Context 2-5 mm. thick, pallid ochre or isabelline, of woven hyphae; skeletal hyphae to 4 μ thick, lumen 1 μ or almost obliterated, unbranched, aseptate, staining deeply; generative hyphee 2-3 μ thick, delicate walled, branched, septate, hyaline, not staining. Pores round, obscurely stratose, 2-4 mm. deep in each layer, pallid ochre in section, 100-200 μ diameter, or 5-6 per mm.; dissepiments 50-150 μ, most 50-75 μ, apices velutinate, enlarged, partly blocking pores. Basidial type clavate, basidia clavate, to 16 x 8 μ, soon collapsing. Spores elliptical, 6-8 x 4-5 μ, smooth, hyaline.
Habitat: Growing solitary, sometimes imbricate, upon bark or decorticated wood of dead standing trunks, associated with a white rot.
Distribution: Australia; Tasmania; New Zealand.
Notes: Resembling F. marginata in many micro-features the species differs in the smaller pores, differently shaped spores, thinner skeletal hyphae and different shape and surface of the pileus. Plants sometimes exude large drops from the surface of the hymenium. Sometimes these persist for so long that pore tissue grows between them, consequently when drops have disappeared the hymenium exhibits a pitted appearance.
In a former paper (1927, p. 210) I listed F. cuneata as a synonym of F. hemitephra, since Lloyd’s scanty description—based on New Zealand specimens at Kew collected from Nothofagus solandri by W.Colenso—suggested that it belonged to this species. His description of the spores was obviously based on those of some contaminating mould. Lloyd’s photograph of a Tasmanian specimen (1924, p. 1329), shows it to be valid and the same as the collections listed above. His Polyporus suaderis appears to be based on the same plant.
From F. hemitephra it may be separated by the different shape and surface of the pileus, larger spores, and absence of an orange zone beneath the cuticle.
|