|
 Inonotus chondromyelus Inonotus chondromyelus
SynonymsInonotus rheades
Pseudoinonotus chondromyelus
BiostatusPresent in region - Indigenous. Non endemic
Images (click to enlarge)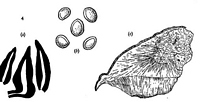
Caption: Fig. 4. I. chondromyleus. a, Setae, x 1000; b, spores, x 1000; c, vertical section through sporophore, x 1. | 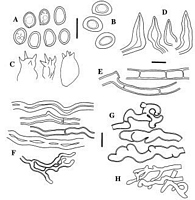
Caption: Fig. 7 Inonotus chondromyelus: A, basidiospores from tubes; B, basidiospores from pileus
surface; C, basidia; D hymenial setae; E, hyphae from near pileus surface; F, hyphae from
context; G, inflated convoluted hyphae of granular core; H bindin | 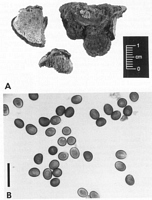
Caption: Fig. 8 Inonotus chondromyelus: A, habit; B, basidiospores from pileus surface (PDD
66487). B, bar = 20 µm. |
Article: Buchanan, P.K.; Ryvarden, L. (1998). New Zealand polypore fungi (Aphyllophorales): three new species and a new record. New Zealand Journal of Botany 36(2): 219-231 (http://www.rsnz.org/publish/abstracts.php).
Description: BASIDIOCARPS probably annual, triquetrous to dimidiate, sometimes lobed; to 4.5 cm
across, 4 cm radius and 2.5 cm thick; margin sharp; pileus surface glabrous or scrupose,
concentrically zonate, orange-brown (58.m Br- 55.s Br) with several to few bands of dark
grey; tubes to 9 mm long, yellow-brown to greyish-brown (76.1y Br- 80.gy.y Br), darker
than context, non-stratose; pore surface pale brown (76.1y Br), discolouring with age to
brown (75.deep.y Br), pores 2-5 per mm, mouths even or sometimes irregularly drawn out;
context bright yellow-brown (72.d.OY - 74.s.y Br), paler than tubes, fibrous, to 10 mm
thick, bounded above by a narrow dark band beyond which is a contrasting reddish brown
tomentum; granular core basally present in context (though perhaps difficult to
discriminate), arising towards point of attachment, with context tissue appearing slightly
darker and denser and sometimes weakly flecked with spots of paler tissue. HYPHAL
SYSTEM monomitic, though with branched hyphae resembling binding hyphae; generative
hyphae brown, in context 3.5-8.5 µm diam., thick-walled to almost solid, wall thickened to
2.5 µm, simple septate with septa widely spaced, straight and mostly sparingly branched
but with occasional narrower side-branches contorted and branched terminally; in
granular core hyphae often wavy to contorted with convoluted and inflated apices to
12.5(-19.5) µm across, with walls thickened and brown, also with narrower, hyaline to
brown, frequently branched hyphae 2.5-3.5 µm diam.; in trama hyphae straight, narrower
than in context, 3-5.5 ltm diam. with wall thickened to 1.5 µm; at pileus surface hyphae
as in context but with more frequent septa; setal hyphae absent. HYMENIAL SETAE
brown, short ventricose with straight pointed apex, thick-walled except in newly formed
setae near tube mouth, embedded in hymenium or projecting slightly beyond, 16-30 x
7.5-13.5 µm. BASIDIA subglobose to broadly ellipsoid, sometimes constricted towards
apical region, 4-sterigmate (sterigmata to 3.5 µm long), septation at base not seen, 14-16
x 8-10.5 µm, typically viewed in preparations as either immature or collapsed during or
following spore development. BASIDIOSPORES oval to broadly ellipsoid; in actively
sporulating tubes spores hyaline, smooth, apiculus small or not evident, IKI-, with wall
thickened to 1 µm, 6.7-9.2 x 5-6.3 µm; spores on pileus surface of some collections and
in tubes of overmature pilel hyaline to light brown, mostly with a single large guttule and
with wall thickened to 2 µm, 7.5-10.5 x 5.5-7.5(-8.5) µm. WOOD ROT not recorded.
Habitat: SUBSTRATA: Known in New Zealand only from dead wood of Nothofagus solandri var.
cliffortioides.
Distribution: New Zealand: Mid Canterbury. Australia: Victoria, South Australia.
Notes: NOTES: This species is newly recorded from New Zealand. Walters (1955, as Inonotus
dryadeus (Pers.: Fr.) Murrill) and Pegler (1964a) provided full descriptions, reporting
conspicuous features including the granular core at the base of the context, presence of
setae, and thick-walled spores. In New Zealand collections the basal granular core is
easily over-looked macroscopically, but differences in hyphal microstructure of the core
and context are evident when comparing microscope preparations of these tissues.
Australian collections (PDD 12402, 13215, 16485, 16486, 20151) show a prominent core,
and pilei are much larger, to 11 cm across, 7.5 cm radius, and 7 cm thick.
The thickness and colour of spore walls appear to gradually change with age of spores;
younger spores present in tubes are hyaline and noticeably thinner walled (Fig. 7A) than
older spores from the pileus surface or from overmature pilei (Fig. 7B, 8B), which varied
in wall colour from hyaline to light brown.
Although the context has a layer separating it from the pileus surface, a feature diagnostic
of Cyclomyces Fr., we follow Ryvarden (1991) and consider the species to belong in
Inonotus P. Karst. on account of its robust fruit-body and large spores. The narrow,
contorted, branched generative hyphae in the context are reminiscent of coltricioid hyphal
construction as described by Corner (1991, fig. 11 right, fig. 30).
In Australia, where this species is known as the Austral Dryad, pilei are reported to
develop from large swellings high up on trunks of Eucalyptus spp., often several pilei
growing together. When fresh, pilei are of light weight and in active growth exude
reddish droplets from the upper surface (Marks et al. 1982).
Inonotus chondromyelus is reported to be common in Victoria and South Australia
(Walters 1955), but is known in New Zealand from only two collections. Along with
several Australian collections, Cunningham (1965) mistakenly assigned his single New
Zealand collection to I. rheades (Pers.) P.Karst. (as (Pers.) Bondartsev & Singer). I.
rheades is a Northern Hemisphere species, restricted to Populus spp., which resembles I.
chondromyelus in having granular core and a similar hyphal structure but differs in
lacking setae and in spore size, 5-6 x 3.5-4 µm (Ryvarden & Gilbertson 1993). From I.
dryadeus, the name earlier used in Australia for specimens of I. chondromyelus (e.g.,
Walters 1955), the latter differs in the presence of a granular core ellipsoid rather than
subglobose spores, and straight not curved setae (Pegler 1964a).
Reid (1967) corrected Cunningham's (1965) mistaken synonymy of the names Polyporus
albertii Lloyd (= Phaeolus albertinii (Lloyd) Reid) and Polyporus victoriensis Lloyd (= I.
victoriensis (Lloyd) Pegler) under I. rheades; both are distinct species. Reid also
expressed doubt that I. rheades occurs in Australasia, but in error cited the Australian
species I. pirisporus Pegler as the correct name for collections labelled I. rheades by
Cunningham. I. pirisporus is distinguished by its apiculate spores and absence of setae
and of a granular core (Pegler 1964a, 1964b), and has not been recorded from New
Zealand. I. victoriensis, described from Australia, differs in lack of a granular core, much
larger pilei than those of I. chondromyelus, a loose hyphal structure in the context, and a
well differentiated pilear crust (Pegler 1964b).
The only recorded New Zealand host, Nothofagus solandri var. cliffortioides, is also host
to a common Inonotus species, I. nothofagi G.Cunn. (Cunningham 1965) which in the
field might be confused with I. chondromyelus. I. nothofagi can be distinguished by the
typically radially striate, often confluent pilei, absence of a granular core, and smaller,
brown spores, 4.7-5.5 x 3.4-4.4 µm, with spore wall thickened to only 0.5 µm. Among 22
New Zealand collections labelled I. nothofagi in PDD, no collections of I. chondromyelus
were found.
Article: Pegler, D.N. (1964). New species of Inonotus (Polyporaceae). Transactions of the British Mycological Society 47(2): 167-173.
Description: Pileus annual, sessile, 4-5.5 X 3-5 X 3-4.5 cm., dimidiate, ungulate, triquetrous in section,
often imbricate or laterally confluent; at first `Dresden Brown', [Colour terms in quotes are
from Ridgway, Color Standards and Color Nomenclature, 1912] `Tawny', or ferruginous,
glabrous, soon developing a hard crust, becoming rimose, and at times radiately rugose
especially towards the margin. Margin acute or obtuse, entire, undulate. Context up to 3 cm.
thick, though thinner towards the margin. Characterized by a large core, situated at the base
of the pileus; this may occupy a considerable proportion of the total volume, is of a granular
nature, permeated throughout by white or yellowish flecks, and contains numerous
conducting organs with yellowish or brownish contents. The remainder of the context which
radiates out from the core is concentrically zoned, fibrous in texture, though watery and
spongy when fresh, but soft and brittle when dry, and with a silky sheen. Colour varies from
'Buckthorn Brown' to `Tawny', but `Ochraceous Buff' or paler towards the pileus surface.
Pore surface convex, often becoming concave towards the margin, ferruginous to dark
umbrinous, narrowly sterile at the margin, usually covered by a cream-coloured pruina
formed of secondary hyphae. Pores medium to large, irregular, round or angular, often
becoming torn, 2-4 per mm., 170-620 µ diam., even. Tubes non-stratified, distinct from the
context, ferruginous, but internally grey, brittle when dry, 2-14 mm. long; dissepiments 25-75
µ thick. Spores, few seen, 7.5-10 x 5.5-7 (8.5 x 6) µ, broadly ellipsoic; sometimes unilaterally
flattened, with a small, lateral apiculus, smooth with a distinctly thickened, colourless or
faintly brownish wall, occasionalh some found embedded within the tube layer or adhering to
the pileus surface making it appear fulvous. Basidia large, colourless, 12-15 x 7-8 µ with 4
sterigmata. Setae present, not abundant, but occasionally localized in small groups, very
thick-walled, but always with a lumen, dark chestnut-brown, appearing black in 10 %
potassium hydroxide solution, 23-35 x 7-10 (-16) µ, ventricose to subulate. Setal hyphae
absent. Context hyphae not agglutinated, dark ferruginous, with a thickened wall but a wide
lumen, septate, branched, 4-10.5 µ, mostly about 5.5 µ diam. Tramal hyphae, tightly
interwoven, not agglutinated, fulvous to ferruginous, very thin-walled, septate, rarely
branched, 1.5-5 µ diam.
Notes: This Australian species has hitherto passed under the name Inonotus dryadeus (Pers. ex Fr.) Murr. [as Polyporus dryadeus]. Cleland (1935),. following the advice given by Lloyd, used this name, though at the same time he suggested that his material might possibly not be the same as the European fungus. When a careful comparison is made of the Australian and European forms it becomes clear that a number of fundamental differences do occur, and that two species are involved though they are enough alike for the confusion in the past to be understandable. Both have a well-developed crust; the spores are large and colourless or nearly so; both have thick-walled, dark-staining setae; and apparently both are prone to exude watery droplets from the pileus surface.
The sporophore of I. chondromyelus, though often imbricate or laterally confluent, consists essentially of a medium-sized, ungulate pileus, which fruits high up on the trunk of the host, and this contrasts sharply with the large, more applanate habit of L. dryadeus, which fruits at or just above ground level. The spores of the former species, though similar in having a thickened wall, are broadly ellipsoid (8.5 x 6 µ) with a lateral apiculus, whereas those of L. dryadeus are subglobose (7.5 x 6.8 µ). The setae, too, are quite different in form, being straight and not curved with a larg, bulbous base. It is in the context that the most obvious distinctions are to be found: L. chondromyelus possesses a granular core, found at the base of the pileus because this is the first-formed structure of the fructification, and even though it may not be well developed it is nevertheless a constant feature which determines the thickness of the sporophore. In the remainder of the context the hyphae have thickened walls and a correspondinly narrower lumen. L. dryadeus never develops a core and the context is composed of thin-walled hyphae with a wide lumen. Other differences may be noted in the pore size and in the nature of the margin.
This species is known only from Australia, where it appears to be not uncommon and is apparently restricted to trunks of Eucalyptus (Walters,1955).
|