|
 Hypoderma rubi Hypoderma rubi
SynonymsHysterium rubi
Lophodermium rubi
Lophoderma rubi
BiostatusPresent in region - Indigenous. Non endemic
Images (click to enlarge)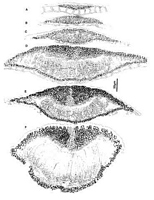
Caption: Fig. 10 Hypoderma rubi (PDD 48949): A-F, unopened ascomata in vertical section, showing
different stages in development. For explanation see notes under H. rubi. | 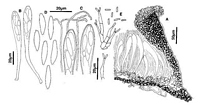
Caption: Fig. 11 Hypoderma rubi: A, margin of ascoma in vertical section (PDD48949). B, asci
(PDD48949). C, apex of asci and paraphyses (PDD 48949). D, released ascospores (PDD
48949). E, conidiogenous cells and conidia (PDD 42790). | 
Caption: Fig. 23 Hypoderma rubi (PDD 42790). Macroscopic appearance of ascomata (x15). | 
Caption: scale = 0.5mm
Owner: J.A. Cooper | |
Article: Johnston, P.R. (1990). Rhytismataceae in New Zealand 3. The genus Hypoderma. New Zealand Journal of Botany 28(2): 159-183 (http://www.rsnz.org/publish/abstracts.php).
Description: Ascomata and conidiomata developing on fallen leaves, in discrete groups within pale, whitish
o yellowish areas of leaf. This paler area rarely associated with narrow, black zone lines. In
surface view ascomata 0.8-3.0 x 0.3-0.6 mm, elliptic to sublinear in outline, tapering to more
or less acute ends. Unopened ascomata with shiny, black walls, sometimes with faint, paler
zone developing along the future line of opening shortly before the ascomata open. Ascomata
open by a single slit lined with a narrow yellow, red-brown, or dark brown zone. Conidiomata
0.1-0.3 mm diam., circular in outline, pale brown at first, becoming dark brown, black around
outside edge and around the central ostiole.
Ascomata subcuticular. In vertical section upper wall of unopened ascomata up to 50-80
µm thick, comprising mostly brown to dark brown, thick-walled, angular cells, but near
middle of the ascomata, along the future line of opening, the inside part of the wall comprises
thinner-walled, paler cells. Wall starts to split open from outside edge, and as the split forms,
hyaline, cylindric cells start to develop along the exposed face of the broken wall. In some
hosts the split in the ascomatal wall may begin to develop prior to the covering host cuticle
breaking. In opened ascomata the upper wall is 60-110 µm thick near the ascomatal opening,
then becoming either abruptly, or gradually, thinner toward the outer edge, where it is 10-25
µm thick. Upper wall comprising dark brown, thick-walled, angular, 4-8 µm diam. cells.
Exposed face of the broken upper wall is lined with cylindric, hyaline, thin-walled, 20-30 x 3-4
µm cells. Lower wall 10-15 µm thick, of 2-4 layers of dark brown, thick-walled, angular to
globose,4-6 µm diam. cells.
Paraphyses 1.0-1.5 µm diam., circinate at apex, extending 5-15 µm beyond asci. Asci
110-160 x 11-14 µm, clavate-stipitate, tapering to small, truncate apex, wall often with small
pore at apex, 8-spored, spores confined to upper half of ascus. Ascospores 15-28 x 2.5-5.5
µm (see notes below), in face view more or less naviculate in shape, widest point near centre,
tapering slightly to rounded apex, and more sharply to acute base. In side view spores slightly
curved. Spores surrounded by well-developed gelatinous sheath. Conidiomata subcuticular.
In vertical section more or less globose in shape, upper wall 5 µm thick, comprising dark
material with no obvious cellular structure, lower wall of 2-3 layers of dark brown, thick-walled, angular cells on which the conidiogenous layer develops. Conidiogenous cells 7-12 x
2-4.5 µm, tapering to apex, proliferating either sympodially, or rarely percurrently. Conidia 3-5 x 1-1.5 µm, cylindric, ends rounded, hyaline, 0-septate.
CHARACTERISTICS IN CULTURE: Spores germinated on agar plates within 24 hours.
Cultures on OA 85 mm diam. after 6 weeks, aerial mycelium white to grey, sparse, agar
surface pinkish to vinaceous brown, with scattered globose, black-walled conidiomata.
Conidiomata splitting to expose grey conidial ooze. Conidiogenous cells and conidia as
described from plant material.
Habitat: Dead leaves, and sometimes twigs, of Aristotelia serrata, Brachyglottis repanda,
Coprosma lucida, C. robusta, Corokia buddleioides, Geniostoma ligustrifolium, Griselinia
littoralis, G. lucida, Leycesteria formosa, Litsea calicaris, Melicytus ramiflorus,
Microsorium diversifolium, Myrsine salicina, Nothofagus solandri var. cliffortioides, N.
truncata, Olearia furfuracea, 0. rani, Pseudopanax arboreus, P. colensoi var. ternatus, P.
crassifolius, Pseudowintera colorata, Quintinia acutifolia, Rhopalostylis sapida, Rubus
cissoides, Rubus fruticosus agg., and Schefflera digitata.
Distribution: Northland, Auckland, Coromandel, Taupo, Taranaki, Wanganui, Wellington,
Nelson, Marlborough, Buller, Westland, North Canterbury, Mid Canterbury, Dunedin,
Fiordland, Stewart I.
Notes: Despite small differences, there is a strong overall similarity between the New
Zealand collections and recent descriptions of H. rubi by Cannon & Minter (1986) and Powell
(1974). The New Zealand collections show a wide range of ascospore length and width (15-28 x 2.5-5.5 µm), and also considerable variation between collections in average spore length
and width. This variation correlates to some degree with host substrate. Ascospores taken
from seven collections on Rubus spp. (both native and introduced) had a range of spore
lengths confined to the upper end of the range for H. rubi as a whole, and spore widths
confined to the lower end of the range of variation, with an average spore size of 24.1 x 3.3
µm. Spores from collections from native hosts, such as Pseudopanax spp. (16 collections,
average spore size 19.6 x 3.9 µm), Griselinia spp. (6 collections, average spore size 19.5 x
3.8 µm), Olearia spp. (6 collections, average spore size 19.0 x 3.8 µm), and Coprosma spp.
(6 collections, average spore size 19.4 x 4.3 µm), had spore lengths confined to the lower end
of the range for H. rubi as a whole, and spore widths confined to the upper end of the range.
Although there is overlap between the range of spore sizes from Rubus versus those from
other hosts, both the ranges and the averages do differ. In all other characteristics of both
teleomorph and anamorph the two groups cannot be distinguished. They are not recognized
here as taxonomically distinct, however the differences in ascospore size may indicate that
they represent genetically isolated populations.
Two distinct species, H. cordylines and H. dundasicum, very similar to H. rubi, have
been recognised from New Zealand, primarily on the basis of ascospore shape and host
preference. All the collections recognised here as belonging in H. rubi have more or less
naviculate ascospores. The spores are widest near the centre, taper slightly to the rounded
apex, and more markedly to the acute base. The ascospores of H. dundasicum are slightly
narrower than those of H. rubi, are cylindric in shape, and although acute to the base, taper
from only very close to the base itself. This character, which correlates with asci slightly
narrower (8-11 µm wide) than those of H. rubi, has been seen in all collections on Ericaceae
from New Zealand.
H. cordylines can be distinguished from H. rubi by ascospore shape, having elliptic or
oblong-elliptic spores which are uniform in shape to both ends of the spore, and in having
longer anamorph conidia. These characters have been seen in all collections examined from
Cordyline and Phormium.
Fig. 10 A-F and Fig. 11 A show stages in ascomatal development of H. rubi. A similar
pattern of development is found in all the Hypoderma spp. included in this paper. When first
visible the developing ascomata comprise a lenticular shaped group of hyaline,
plectenchymatous cells beneath which is the lower wall layer, comprising a single row of
brown, thick-walled cells (Fig. 10 A). The lower wall under-goes more or less no change
during subsequent stages of ascomatal development. At about the same time as the upper wall
of the ascomata develops, a cavity forms within the hyaline plectenchyma, and paraphysis-like
elements extend up into this cavity (Fig. 10 B, C). As the ascomata develop they increase in
size, the upper wall becomes thicker toward the centre of the ascomata, the wall becomes
internally differentiated, the paraphyses become longer, and asci begin to develop (Fig. 10 B,
C, D). The upper wall comprises mostly brown, thick-walled, angular cells, but near the
centre of the ascomata, in the inner half of the wall, is a wedge-shaped group of pale, thin-walled cells, and lining the inner edge of the wall is a layer of hyaline, short-cylindric,
periphysoid-like cells (Fig. 10 D, E). The ascomatal upper wall starts to split open from the
outside, and at the same time as the split develops, hyaline, thin-walled, cylindric cells form
along the exposed face of the broken wall (Fig. 10 F), and in opened ascomata form a well-developed, persistent layer (Fig. 11 A). The periphysoid-like cells evident in immature
ascomata are not visible in opened ascomata, presumably crushed by pressure exerted by the
developing hymenium.
|