|
 Hamaspora australis Hamaspora australis
SynonymsHamaspora acutissima
BiostatusPresent in region - Indigenous. Non endemic
Images (click to enlarge)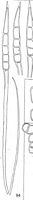
Caption: FIG. 94. Hamaspora acutissima Syd. Teleutospores from Rubus australis Forst. f. | 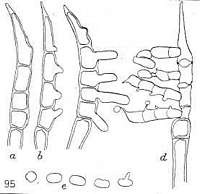
Caption: FIG. 95. Hamaspora acutissima Syd. Teleutospores, showing different stages during
germination; (e) basidiospores, one germinating. | 
Caption: FIG. 4. Hamaspora acutissima Syd. Teleutosori on Rubus australis Forst. f. Note the long
and much-entwined fibrils. Arrow points to sori from which the fibrils have disappeared. | 
Owner: Herb. PDD | 
Owner: J.A. Cooper |
Article: Gadgil, P.D. (in association with Dick, M.A.; Hood, I.A.; Pennycook, S.R.) (2005). Fungi on trees and shrubs in New Zealand. Fungi of New Zealand. Ngā Harore o Aotearoa 4: xi + 437 p. Hong Kong: Fungal Diversity Press.
Description: Type: Rust and Smut Fungi; Description: Telia crowded in irregular groups, elliptical, brown, powdery, up to 3 mm in diameter; on the lower surfaces of leaves, in pale yellow indefinite spots coincident on both leaf surfaces. Teliospores aggregated into pale yellow to white persistent filaments up to 2 cm long, long-cylindrical, 3–5-septate, apex tapering to a fine, sharp point, 100–180 × 14–22 μm; pedicel persistent, up to 600 om long.
Distribution: Distribution: Bay of Plenty, Taupo, Taranaki, Wanganui, Wellington, Gisborne, Hawkes Bay, Wairarapa, Nelson, Buller, Westland, Marlborough Sounds, North Canterbury, Mid Canterbury, South Canterbury, Otago Lakes, Central Otago, Dunedin, Southland, Stewart Island.; 1st Record: Cunningham (1924a: as Hamaspora acutissima).
Article: Cunningham, G.H. (1924). The Uredinales, or rust-fungi, of New Zealand: supplement to Part 1; and Part 2. Transactions and Proceedings of the New Zealand Institute 55: 1-58 Wellington:.
Description: 0. Spermogones immersed, hypophyllous, associated with the uredosori.
II. Uredosori hypophyllous, scattered, orbicular, 0.25-0.5 mm. diam., golden-yellow;
surrounded by and mixed with numerous incurved hyaline cylindrical paraphyses. Spores
globose, obovate, or shortly elliptical, 20-25 X 17-19 mmm.; epispore hyaline, finely and
densely verruculose, 2 mmm. thick, thickened at the apex to 3-5 mmm., cell-contents pallid
yellow; germ-pores scattered, 6-8, obscure.
III. Teleutosori hypophyllous, scattered or crowded in irregular groups, seated on indefinite
pallid spots which are visible on the upper surface, elliptical, 0.5-1 mm. diam., pulverulent,
surrounded by the ruptured epidermis. Spores aggregated into pallid-yellow fibrils up to 20
mm. long, fading with age, 4-6-celled, long-cylindrical, 100-180 X 14-22 mmm.; apex
strongly acuminate, tapering to a fine sharp point, thickened up to 10 mmm., base truncate;
not constricted at the septa; epispore hyaline, smooth, 1 mmm. thick, cell-contents orange,
granular; pedicel persistent, continuous with the spore, tapering basally to a fine point, up to
600 mmm. long, 10-15 mmm. thick immediately below the spore, hollow; germ-pore solitary
in each cell, indistinct.
Distribution: Distribution : Java; Queensland.
Notes: The host is endemic, and is common throughout. (Cheeseman, 1906, p. 125.) The
teleutospore stage only has been collected in New Zealand; this is common, and is readily
observed owing to the conspicuous nature of the spore-masses.
The description of the uredospores given above is drawn up from information kindly
supplied by. Mr. C. C. Brittlebank, Melbourne.
The spores and their pedicels are inextricably interwoven into filaments, so that it is
difficult to separate out single spores for the purposes of measurement and drawing. These
filaments are formed in the following manner: The sori are at first bullate and covered by
the epidermis; this soon becomes ruptured, owing to the increasing pressure exerted by the
developing spores, and the spores become exposed, when they are seen as a pulvinate mass
standing above the leaf-surface about 0-5 mm. Spores are being produced from the same
sorus during the whole of the summer months, and appear to be developed in batches. As
each successive batch develops, the spores become forced between the pedicels of the
preceding batch, which are consequently: carried upward away from the base of the sorus.
Gradually the filament becomes formed, and, as the spores and pedicels of successive
batches are closely interwoven, the filament does not break up, but remains compact, and
may persist for several seasons, especially if it happens to become entangled with the
spines which cover the undersurface of the leaf. The filaments do not, as a rule, remain for
any length of time attached to the sori, so that they may frequently be seen on the same leaf
with the detached filaments. The empty sorus usually contains numerous incurved,
cylindrical paraphyses. Germination occurs as soon as the spores mature, and, as spores of
all ages usually occur in the same filament, different stages of development may readily be
obtained. The first indication of germination is the appearance of a slight swelling on one
side of a cell; this is followed by the gradual development of the basidium, which protrudes
at this point. At first unicellular, the basidium soon becomes four-celled by the appearance
of transverse septa. Shortly after the septa appear the sterigmata grow out, one from each
cell of the basidium; on each a small, smooth-walled, colourless basidiospore appears. The
basidia are allantoid, and may attain a size of 60 X 10 mmm.; the basidiospores are obovate
or reniform, and usually 15 X 10 mmm.
Until 1912, when P. and H. Sydow (l.c.) separated the species described above, one species
only was known. This was by Thuemen in 1875 (Flora, vol. 58, p. 379) described as
Phragmidium longissimum. Two years later Koernicke (l.c.) placed it in Hamaspora, a
genus he erected to contain it and another form with similar spores (now
Gymnosporangium Ellisii Farl.). In 1888 De Toni (in Sacc., Syll., vol. 7, p. 750) compiled
it under Phragmidium. Massee in 1893 (Grey., vol. 22, p. 17) considered that it was neither
Hamaspora nor Phragmidium, but stated that he did not care to undertake the
responsibility of forming a new genus (!). Dietel (1900, p. 73) considered that there were
no grounds for separating it from Phragmidium, although he recognized a resemblance to
Gymnosporangium in the teleutospore structure. McAlpine (1906, p. 187) in 1906 included
it under Phragmidium, but mentioned the fact that it differed considerably from other
species included in that genus.
|