|
 Gloiocephala rubescens Gloiocephala rubescens
SynonymsCampanella rubescens
BiostatusPresent in region - Indigenous. Non endemic
Images (click to enlarge)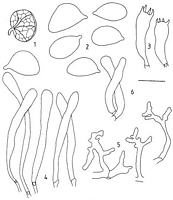
Caption: C. rubescens. 1. Basidiomes (2x): 2, spores;
3, basidia: 4, cheilocystidia; 5, pileipellis elements; 6, caulocystidia. | 
Caption: ZT8739
Owner: E. Horak: © Creative Commons Attribution-Noncommercial 3.0 New Zealand | 
Caption: Dried type specimen
Owner: Herb PDD | 
Caption: ZT0522
Owner: E. Horak: © Creative Commons Attribution-Noncommercial 3.0 New Zealand | 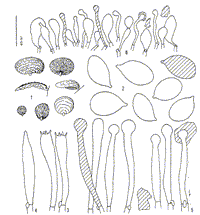
Caption: Fig. 1 (1-6). Gloiocephala rubescens (Segedin) Desjardin & E. Horak (ZT 522).- 1. Basidiomes.-
2. Basidiospores.- 3. Basidia.- 4. Basidiole.- 5. Cheilogloeocystidia.- 6. Pileipellis (x500). |
Article: Segedin, B.P. (1993). Studies in the Agaricales of New Zealand: some new and revised species of Campanella (Tricholomataceae: Collybieae). New Zealand Journal of Botany 31: 375-384 (http://www.rsnz.org/publish/abstracts.php).
Description: Basidiome 12 x 9 mm diam., orbicular to reniform,
fleshy, fragile, easily detached from substratum, stipitate. Pileus pinkish
cream, "chamois to cinnamon buff", becoming red with age, gelatinous,
surface viscid, tessellate corresponding with lamellae. Hymenophore white to
pinkish buff at first but becoming crimson to rusty red, "orange cinnamon
to mikado brown, deep corinthian red to acajou red" (colours in quotation
marks from Ridgway (1912), fide R. H, Petersen pers. comm.), staining appearing
first at the margins of the long lamellae, strongly intervenose, with a small
number (3-5) of long lamellae reaching the stipe, and between them secondary
lamellae anastomosing at a lower level. During drying, the margins of lamellae
exude a sticky-looking red substance. Stipe 2 x 1 mm, excentric to mainly lateral,
cylindric, solid, pink, pubescent. Smell and taste unknown. Spore print white,
copious.
Spores 10.5-14.5 x 6.5-9.0 (13.0 x 7.5) µm, Q =
1.73, elliptical in face view, flattened on adaxial side and often conspicuously
humped on abaxial side in lateral view, hyaline, inamyloid, not dextrinoid,
acyanophilic, thin-walled, some with refractive globules; contents of spores
discolour ferruginous red with age. Basidia 35-45 x 5-12 µm, elongate-clavate,
with 4 sterigmata, up to 6 µm long, contents yellow granular when young. Cheilocystidia
70-100 x 3-9 µm, cylindrical to narrowly clavate, thin-walled, contents becoming
reddish with age, or sometimes with red material forming a cap, crowded, forming
a wide, sterile, lamellar margin. Pleurocystidia none but some fusoid basidioles
present. Trama irregular, gelatinised, some narrow, oleiferous hyphae present.
Context subgelatinous, of narrow (3-5 µm) hyphae, loosely woven with very distinct
clamp connections; some narrow, oleiferous hyphae ending in the pileipellis
as clavate cells with pink contents. Pileipellis of repent hyphae giving rise
to narrow diverticulate hyphae in a loose Rameales structure. Caulocystidia
40-50 x 6-8 µm, cylindrical to slightly clavate, frequent, very similar to cheilocystidia.
Habitat: On decaying nikau palm fronds (Rhopalostylis
sapida Wendi. & Drude, an indigenous palm), in mixed podocarp dicotyledonous
forest.
Notes: This species bears a striking resemblance
to C. pustulata (Berk.& Br.) Pegler, another red-staining species,
first recorded by Berkeley & Broome (1873) growing on decaying palm fronds
in Sri Lanka, and described again by Petch (1910) and Pegler (1986), also from
Sri Lanka. Cooke (1892) recorded it for Australia but Petch (1910) stated that
"Cooke's figure in the Handbook of Australian fungi" (which shows
a very small Campanella-like fungus entirely ochraceous in colour) "has
no relation to this plant at all", a view subscribed to by Lloyd (1919).
The New Zealand species is like the Sri Lankan in certain respects such as the
substratum, the colour changes of the basidiomes, the similar appearance of
the extruded, glutinous substance from the lamellar margin in dried material,
and the ferruginous staining character of the spores and cheilocystidia, but
it differs in that the basidiomes are considerably larger than those in the
Sri Lankan type material examined and are laterally stipitate instead of being
attached by a pseudostipe (Pegler 1986), and the cheilocystidia are longer and
narrower than those figured by Pegler. Pegler did not report caulocystidia or
oleiferous hyphae in C. pustulata. It was difficult to identify the tissues
of the type with any precision because of the small amount and size of the material
available, but the spores of C. pustulata appear to be similar in size
but slightly more spherical (Q = 1.3 instead of 1.7) than those of C. rubescens.
On the basis of the large spore size, both these species would fit in Singer's
(1975) subsection Gigantosporae, type species C. gigantospora Singer
from Australia, but neither species has the diverticulate to somewhat thick-walled
cystidia that were confirmed from an examination of the type of C. gigantospora.
C. rubescens appears to be fairly common in New Zealand, on decaying fronds of the indigenous
nikau palm (R. H. Petersen pers. comm.). The red staining of the basidiome is
a very distinctive character, making it easy to recognise.
Article: Desjardin, D.E.; Horak, E. (1997). Marasmius and Gloiocephala in the South Pacific Region: Papua New Guinea, New Caledonia, and New Zealand taxa. Bibliotheca Mycologica 168: 152 p.
Description: Pi1eus 5-25 mm, polymorphic, orbicular, fan-shaped, ear-shaped or conchiform; margin incurved;
surface covered with conspicuous, concentric, tesselate ridges, dull, moist to dry, glabrous; texture
strongly gelatinous-cartilaginous, tough; at first whitish to pale-ochre, slowly changing to deep
reddish brown or copper brown in age. Context pale beige to pale ochre, strongly gelatinous, car-tilaginous.- Lamellae narrowly adnate, distant (3-8) with 2-4 series of lamellulae, strongly
anastomosing or interconnected, transverse lamellulae often at a lower level, sometimes merulioid;
white to off-white, becoming pinkish buff to pinkish cinnamon, eventually becoming concolorous
with pileus surface; edges even, darker red or brown.- Stipe 1-3 x 1 mm, lateral or eccentric, rarely
absent, cylindrical, equal, solid, conspicuously pruinose, insititious; concolorous with pileus.- Odor
and taste not distinctive.
Basidiospores 10.0-14.5 x 6.5-8.0(-9.0) µm, elliptical to ovoid, with broad lateral apiculus, smooth,
hyaline, often with strongly refractive red-brown globules, inamyloid, thin-walled.- Basidia 45-50 x
7-8 µm, elongate clavate to subcylindrical, 4-spored, clamped.- Basidio1es fusoid.-
Chei1og1oeocystidia abundant (lamellar edge sterile), 40-60 x 4-6 µm, tibiiform to slender fusoid
with capitate apex, capitulum 8-10 µm diam., or, slender clavate and non-capitate, hyaline, thin-walled, often with reddish brown plasmatic or coagulated pigment, upper portion often covered with
reddish brown resinous exudate.-Pleurogloeocystidia absent.- Hymenial metuloids absent.-
Pileipellis hymeniform, composed of Globulares-type cells plus pileogloeocystidia; Globulares-type cells 18-40 x 8-18 µm, clavate to broadly clavate, thin-walled, hyaline with reddish brown
globular contents; pilogloeocystidia scattered, 35-52 x 3.5-5.0 µm, tibiiform, capitulum 6-8 µm
diam., hyaline with reddish brown globular contents; reddish brown exudates adhering to surface of
pileipellis; metuloids absent; diverticulate cells absent.- Pileal and lamellar tissue inamyloid,
gelatinous.- Caulogloeocystidia like cheilogloeocystidia, but smaller.- Clamp connections present.
Habitat: Habit, habitat and distribution.-Solitary, in groups on rotting leaves of Rhopalostylis sapida
(Palmae). New Zealand, Sri Lanka.
Notes: Gloiocephala rubescens was described initially as a species of Campanella (Segedin, 1993)
based on the observation of diverticulate pileipellis elements, gelatinous tissues and
Campanella-like macromorphology. Examination of a paratype specimen collected by R. H.
Petersen from which some of the protologue data were obtained, indicated that the pileipellis is
not composed of a cutis of diverticulate hyphae (i.e., a Rameales-structure typical for
Campanella), but rather is a hymeniform layer of clavate cells plus pileogloeocystidia (the
pileipellis is often obscured by the adherent resinous reddish brown exudates from
pilogloeocystidia and is therefore difficult to analyze). This pileipellis anatomy, in conjunction
with tibiiform hymenial gloeocystidia with pigmented contents is diagnostic for the genus
Gloiocephala and the species is formally transferred accordingly. The strongly anastomosing to
nearly poroid hymenophore, small lateral stipe, color changes from white to reddish with age, and
growth on palm fronds is diagnostic.
Gloiocephala rubescens belongs in sect. Gloiocephala, subsect. Macrosporae where it is most
closely allied with G. palmarum Singer described from Bolivia. Gloiocephala palmarum differs
in forming pilei with orange rather than red pigmentation, lamellae that are less intervenose and
not at all poroid, narrower basidiospores (4.5-6.5 µm broad), and metuloid cystidia 55-135 µm
long in the pileipellis and stipitipellis (fide Singer, 1960).
|