|
 Lentinellus novae-zelandiae Lentinellus novae-zelandiae
SynonymsLentinellus marginatus
Lentinus novae-zelandiae
Pocillaria novae-zelandiae
BiostatusPresent in region - Indigenous. Endemic
Images (click to enlarge)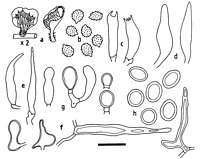
Caption: Fig. 4 L. marginatus. a, basidiomata whole and in section (x 2); b, spores; c, basidia; d, lamellar pseudocystidia; e, lamellar leptocystidia; f, pileal
pseudocystidia; g, stages in chlamydospore formation and secession; h, seceded chlamydospore | 
Caption: Dried type specimen
Owner: Herb PDD | 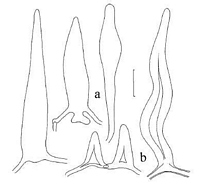
Caption: Fig. 24. Lentinellus novae-zelandiae, pileicystidia. a. TFB 7104. b. PDD 62527. Standard
bar= 10 µm. |
Article: Segedin, B.P. (1996). A new species of Lentinellus (Hericiales, Lentinellaceae) and a revision of taxa attributed to Lentinellus in New Zealand. New Zealand Journal of Botany 34(2): 249-261 (http://www.rsnz.org/publish/abstracts.php).
Description: Basidioma convex, dimidiate to ligulate, sessile or laterally stipitate. Pileus 10-25 mm diam. (dry measurement), dull pinkish-fawn when fresh (G. M. Taylor
field notes), when dried greyish-orange, sahara, sienna (6C5-6D5 K&W), darker at the margin, surface smooth, sometimes with scattered, minute white to
buff hairs near the stipe attachment, margin inrolled, lacerate. Lamellae pale pinkish-fawn when fresh (G. M. Taylor field notes), with a white margin (J. M.
Dingley field notes), drying yellowish-brown (6C4 K&W), relatively broad, moderately distant, in three series, adnate to decurrent, edge broadly lacerate.
Stipe when present lateral, small, button-like or up to 10 mm long by 3 mm broad, cylindrical, typically clothed in a tangled tomentum through which
protrude dark red-brown to black bristly hairs, up to 2 mm long, particularly noticeable in older basidiomata. Taste and smell unknown. Spore print white,
becoming cream.
Spores 4.0-5.5 x 3.5 4.5 (4.9 x 4.0) µm, Q = 1.2, subglobose, hyaline, wall strongly amyloid, with short peg-like ornamentation and distinct apiculus. Basidia
20 - 30 x 6 µm, clavate, mostly 4-spored, sometimes 2-spored, each with clamp connection on basal septum. Leptocystidia plentiful on all parts of the
lamellae, mostly subulate, narrowly to broadly fusiform or narrowly lageniform, 30-60 x 3 - 12 µm, thin-walled, usually without contents and each with a
clamp connection at the base. A few clavate to lanceolate pseudocystidia also present. Trama of more or less parallel, thin-walled, clamped, descending
hyphae, up to 3 µm diam. Many parallel, oleiferous hyphae present, 3-5 µm diam., becoming contorted, with some empty segments and slight epimembranal
incrustation, near the very narrow, cellular sub-hymenium. No thick-walled hyphae in trama. Trama ochraceous in KOH, negative in Melzer's reagent.
Context of interwoven hyphae, relatively thin-walled, up to 6 µm diam., with plentiful oleiferous hyphae concentrated particularly near the pileipellis.
Oleiferous hyphae up to 7 µm diam., often much contorted and with swellings up to 14 µm diam. Skeletal hyphae absent. Middle region of context
dextrinoid but colour fades fairly quickly. Pileipellis of repent, interwoven, yellowish-brown, thin-walled hyphae, 2 - 3 µm diam., with many interspersed
oleiferous hyphae, the ends of which protrude as numerous pseudocystidia, especially near the stipe attachment. Pseudocystidia clavate, lageniform or
lanceolate, 50- 90 x 12 - 13 µm, walls only slightly thickened at first but those towards the stipe becoming thick-walled. Sections through the margin of the
pileus show clusters of clavate cells 20-30 x 5 - 7µm, intermingled with basidia and pseudocystidia. The apices of these clavate cells become converted into
thick-walled chlamydospores, subpyriform when first produced but after secession becoming subspherical, 5.5-8.0 x 4.5-6.5 (7.15 x 5.8) µm, Q = 1.2, with
walls up to 3 µm thick, inamyloid, not dextrinoid. Stipe of more or less parallel hyphae up to 3 µm diam. with slightly thickened walls, together with
oleiferous hyphae of slightly larger diameter and some thick-walled hyphae up to 6 µm diam., with walls up to 2 µm. Surface tomentum of stipe of tangled,
hyaline, branched, narrow (up to 2 µm) clamped hyphae, mixed with oleiferous hyphal endings and a few fusiform leptocystidia; chlamydospores, detached
from the margin of the pileus, are commonly found adhering to the tomentum hyphae, but never seen with a germ tube. The tough, bristly hairs up to 2 mm
long that protrude through the tomentum consist of narrow (1-2 µm diam.), slightly thick-walled, brown, agglutinated hyphae, with occasional clamp
connections.
Habitat: HABITAT: on standing or fallen rotten wood in a mixed beech/podocarp forest, disturbed, with some adventives.
Notes: ETYMOLOGY: Referring to the chlamydospores on the margin of pileus and the white edge of the lamellae.
The production of chlamydospores on the margin of the cap is a very distinctive character in Lentinellus marginatus. L. cochleatus seems to be the only other
species reported to produce chlamydospores but they are described as being either on the stipe (Miller & Stewart 1971) or formed interstitially beneath the
pileipellis (Breitenbach & Kranzlin 1991). Also, the basidiomata of L. cochleatus are different in form, with well developed, long stipes in tightly fused
caespitose clusters (Miller & Stewart 1971).
Article: Stevenson, G. (1964). The Agaricales of New Zealand: V. Kew Bulletin 19(1): 1-59.
Notes: The type specimen is lost and the description is doubtfully adequate.
Article: Segedin, B.P. (1996). A new species of Lentinellus (Hericiales, Lentinellaceae) and a revision of taxa attributed to Lentinellus in New Zealand. New Zealand Journal of Botany 34(2): 249-261 (http://www.rsnz.org/publish/abstracts.php).
Notes: Shown to be a synonym of Lentinellus ursinus by Pegler 1983.
Berkeley's (1855) description is "Subimbricated. Pileus 1 inch or more long and broad, thin, flabelliform, suborbicular or reniform, bay brown, clothed
behind with short velvety olive down. Stem obsolete. Gills same colour as the pileus, narrow, decurrent behind; edge thin, lacerated. Closely resembling L.
castoreus, of which I have an authentic specimen, but differing in size and narrow gills."
The type material (New Zealand: Colenso, Herb. Phillips, ex Herb. Berkeley, K) consists of 4 basidiomata up to 26 x 17 mm diam. Their characters all
confirm Pegler's identification of the species as yet another synonym of L. ursinus.
Article: Massee, G.E. (1899) [1898]. The fungus flora of New Zealand. Transactions and Proceedings of the New Zealand Institute 31: 282–349 Wellington:.
Description: Sessile, attached by a narrowed base, fan-shaped, reniform or suborbicular, thin and flexible,
bay-brown, clothed behind with short velvety olive down, about 2.5 cm. long and broad; gills
decurrent behind, narrow, edge thin and torn, coloured like the pileus.
Habitat: On dead wood.
Distribution: New Zealand.
Notes: Closely resembling Lentinus castoreus, but smaller, and with narrower gills. There is no
specimen in Berkeley's herbarium.
Article: Petersen, R.H.; Hughes, K.W. (2004). A contribution to a monograph of Lentinellus. Bibliotheca Mycologica 198: 1-179.
Description: Basidiomata gregarious, flabelliform, spathulate, ligulate when young to subdimidiate when
mature, convex to subconchate, sessile to pendent (i.e. not everted-curled like L. cochleatus).
Pileus 7-32 mm broad, up to 24 mm deep, subundulate to delicately radially rivulose, "Rood's
brown" to "pecan brown" (6C5-6135), russet to (where poorly dried) brown-black, "orange
buff' to "capucine buff' (5A5-6) or duller (5B5-7), with spots of "buckthorn brown" (5136),
surface smooth outward, minutely pubescent to minutely micaceous at disc, more hirsute or
hoary at stipe attachment; hoary ornamentation yellow, drying pallid gray; margin
translucent-striate, decurved, fimbriate to crenulate but not striate, darker than pileus, as
though hygrophanous (5B5); flesh thin (less than 1 mm thick). Lamellae adnate,
subdecurrent, subdistant, up to 3.5 mm deep, in at least three ranks, "pale cinnamon pink"
(5A2-3) to "cinnamon buff' when fresh, drying to "tilleul buff'.to (poorly dried) dark brown
(5135, 6C4); margin coarsely digitate-serrate, reported as minutely white. Stipe lateral to
absent, 2-10 X 2-4 mm, prominent, reduced to a knot or absent, more or less equal (when
present), irregular in cross-section (i.e. not terete, fluted by lamellar ridges), near "buckthorn
brown" (5137), dark brown, "drab," to "light drab," strigose, plushy to covered with hyphal
spikes. Odor very weak, typical; taste very slowly weakly acrid, mildly anesthetic to tip of
tongue.
Pileipellis a repent, radial, coherent layer of generative hyphae involved in mucus; hyphae
2.0-7.0 µm diam, hyaline to yellowish brown (BF, KOH), thin-walled, clamped; mucus epi-
and interhyphal, discharged in globules in squash mounts (PhC, KOH), amorphous, not
granular. Suprapellis outward undeveloped but lanceolate pileicystidia emergent from
generative hyphae; pileicystidia (Fig. 24) 55-92 X 6.7-13.0 µm, ranging from elongate-ellipsoid with
bluntly rounded apex to fusoid with prolonged beak, to fusiform-lanceolate,
gradually tapered to bluntly lanceolate tip, hyaline, sometimes biaxial, thin-walled,
inamyloid; inward pileicystidia gloeoplerous, 150-340 X 6.5-17.0 µm, narrowly vermiform to
nematoform, tapering to narrow apex, undulate to irregularly swollen, thick-walled (wall up
to 1.8 µm thick), yellow-refringent (PhC, KOH), inamyloid; contents glassy to finely
coscinoidal; pileileptocystidia numerous, 2.0-3.0 µm diam, hyaline, filamentous, rounded at
apex, twisted to delicately undulate, emergent up to 45 µm. Subpellis of outer pileus dimitic:
1) generative hyphae 3.0-6.0 µm diam, hyaline, thin-walled, clamped; and 2) gloeoplerous
hyphae abundant, sinuous, yellow-refringent (PhC, KOH); contents cosdinoidal; subpellis of
inner pileus dimitic: 1) generative hyphae undulate, thick-walled (wall up to 0.7 µm thick),
hyaline, clamped; and 2) gloeoplerous hyphae occasional, as in outer subpellis.
Pileus trama interwoven, dimitic: 1) generative hyphae 2.0-7.5 µm diam, hyaline, thick-walled
(wall up to 1.3 µm thick, commonly occluding cell lumen), glassy-refringent over
short lengths, long-celled, branched, inamyloid to weakly amyloid (pale blue-gray),
conspicuously and frequently clamped, occasionally ornamented with minute warts on
exterior surface and then appearing minutely banded; and 2) gloeoplerous hyphae common
(not abundant) near surface, less so inward, 3.5-7.0(-14.0) µm diam, yellow-refringent (PhC,
KOH), thin-walled, emergent up to 15 µm; and 4) leptocystidia (Fig. 25c) filamentous, 1.8-3.0 µm diam,
yellow-refringent (PhC, KOH), emergent up to 15 µm; contents coscinoidal.
Basidiospores (Fig. 25d) 4.0-5.7 X (3.5-)4.0-4.5 µm (E = 1.08-1.33; Em =
1.20; Lm = 4.85
gym), ovate to broadly ovate to subglobose, thin-walled, weakly to moderately amyloid;
ornamentation of densely scattered, strongly amyloid prickles barely visible at 1500X;contents homogeneous to uniguttulate.
Habitat: Habitat: on standing or fallen rotten wood in a mixed Nothofagus/Podocarpus forest
Distribution: New
Zealand, Patagonia, Tierra del Fuego.
Notes: Commentary Although centrally to eccentrically stipitate basidiomata of L. singeri are found (see-Horak,
1979), some basidiomata closely resemble those of L. fabelliformis and/or L.
occidentalis (laterally stipitate; pileus broadly cuneiform to subdimidiate). In this feature,
however, basidiomata are not of the everted-curled stature of members of the L. cochleatus
complex.
Neither pleurocystidia nor pileicystidia can be reliably referred to generative or gloeoplerous
hyphal systems. Neither structure could be traced to a gloeoplerous hypha and contents
appeared hyaline and homogeneous under phase contrast microscopy. Horak correctly
illustrated pileicystidia as basally bifurcate, assumedly arising from generative hyphae of the
subpellis or trama. Nonetheless, occasionally cell contents are less than homogeneous, but
subtly multigranular as though gloeoplerous. In L. tridentinus, pileicystidia are definitely
gloeoplerous in color and light refringence, and differ in form (pileicystidia vermiform in L.
tridentinus; hymenial cystidia clavate to bluntly rounded).
A molecular phylogenetic reconstruction places L. singeri as virtually congruent with L.
marginatus ss Segedin (q.v.). The two species share several rather unique characters: 1)
attachment to substratum; 2) small basidiomata; 3) minutely micaceous appearance of inner
pileus surface (30X); 4) similar pileal pseudocystidia; 5) subdistant, coarsely serrate lamellae;
6) relatively large basidiospores. Examination of the few available specimens of each species
lead us to conclude that they are conspecific.
Horak (1979) referred to Lentinellus omphalomorphus (Bertero & Montagne) ss. Singer
as a synonym of L. singeri, but presented no evidence of how "ss. Singer" differed from the
original descriptions and/or specimen by Montagne. Likewise, no evidence was reported
about why the taxon required a new name unless "ss Singer" could be shown not to agree
with the original material.
Nonetheless, L. singeri was given as "sp. n." by Horak, not as "nom. nov." as would have
been required if Agaricus omphalomorphus Bert. apud Mont. had been a later homonym.
Likewise, Singer (1969) recombined Agaricus omphalomorphus in Lentinellus, and while
citing several Singer collections, did not report examination of Montagne's type
specimen. We were unable to examine Montagne's type specimen (PC), so the identity of
Montagne's organism remains in doubt, and the correlation of Singer's sense of the name
cannot be ascertained without examination of all pertinent material. Until this has been
accomplished, we are reluctant to repeat doubtful synonymy.
Although we have examined the specimens which Segedin (1996) cited under L.
marginatus, we have been unable to observe the "chlamydospores" described and figured
by Segedin. Instead, the only structures which could be construed as such were
skeletalized pileicystidia near the pileus margin, elongate-ellipsoid in outline but never
disarticulated. Moreover, Segedin (1996) described L. marginatus even after having
examined the type specimen of Lentinus novaezelandiae, apparently based on the putative
chlamydospores in L. marginatus.
Some basidiomata of L. novaezelandiae are virtually indistinguishable from those of L.
occidentalis. Those of the latter are also characterized by pileicystidia of comparable
dimensions to those of L. novae-zelandiae, and spore statistics are not significantly
different. DNA sequences, moreover, show L. occidentalis to be in a clade with L.
montanus and closely related to L. novae-zelandiae.
Although dimitic, generative hyphae of the pileus trama in L. novae-zelandiae are often
skeletalized and then sometimes faintly amyloid. This construction is quite like that of L.
fabelliformis, basidiomata of which are similar to those of L. novae-zelandiae.
Pileicystidia are absent in L. fabelliformis, however, and the species is known to occur
only in the North Temperate Zone.
|