|
 Rosellinia arcuata Rosellinia arcuata
BiostatusPresent in region - Indigenous. Non endemic
Images (click to enlarge)
Caption: PDD 82750
Owner: P.R. Johnston | 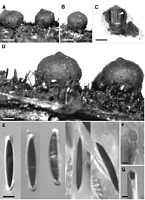
Caption: Fig. 7 Rosellinia arcuata. A, B, D, Stromata, D showing synnemata; C, Vertical section of
stroma, arrow points to collapsed perithecium; E, Asc | 
Caption: Fig. 8 A-D, Rosellinia arcuata, PDD 41969: A, Ascospores; B, Ascus apical ring; C,
Conidiophores and conidia on the host; D, Conidiophores and conidia on OA; E-G, R.
freycinetiae, PDD 20580: E, Ascus a | 
Caption: C-3686
Owner: Herb. PDD | |
Article: Petrini, L.E. (2003). Rosellinia and related genera in New Zealand. New Zealand Journal of Botany 41(1): 71-138 (http://www.rsnz.org/publish/abstracts.php).
Description: Subiculum persistent, brown to dark brown, wiry, appressed, with synnemata. Stromata
(1250)1766 ± 330(2500) µm high, (1125)1740 ± 312(2250) µm wide (n = 30), globose to
subglobose with almost flattened top, cupulate, often with a short cylindrical base immersed
in the subiculum, copper brown, dark brown, black around the ostioles, smooth, solitary to
crowded and laterally compressed. Ostioles finely to coarsely papillate. Ectostroma
(75)100-125 µm thick, black. Entostroma white, confined to base. Perithecia detached and
collapsed in mature material. Ascus apical ring (6.7)8.3 ± 0.8(9.6) µm high, upper width
5.7-6.2 µm, lower width 3.8-5.7 µm (n = 16), J+, dark blue. Ascospores (37.4)48.6 ±
4.2(58.5) µm long, (4.8)7.6 ± 0.8(9.6) µm wide (n = 167), inequilateral, narrowly ellipsoidal
to needle shaped, with narrowly rounded ends, brown to dark brown, with straight, 8-12 µm
long germ slit centred on the flat side, both extremities and flat side surrounded by a slimy
sheath, 2-3 µm thick at ends, 1 µm thick at side. Synnemata up to 2000 µm, conidia 4-5 x 3-4
µm.
Culture on OA after 20 days under 12 h dark and 12 h UV and fluorescent light 7 cm diam.,
centre (3 cm) black from confluent tufts of black hyphae shaped as witches' brooms, otherwise
white, hyphae very short, sterile. On CMD after 43 days under same conditions covering
whole plate (9 cm diam.), black with cottony white margin. Synnemata profusely arising at the
margin and on the original inoculum, 0.5-1 mm long, acute, unbranched, black, without
stromatic base, composed of 3-4 µm wide smooth, brown, branching, septate hyphae, often
sterile. Free conidiophores forming on one area of the original inoculum, long, heavily and
irregularly branched. Conidiogenous cells 18-48 3-4 µm (n = 9), terminal, geniculate, smooth,
subhyaline to pale tantowards the base. Conidia 3.5-5(7) x 2-3.5 µm (n = 22), forming at the
tip of each synnema as loose, white-grey areas, oblong with flat, non-protuberant base bearing
a minute frill being more refractive than the rest of the conidial wall, smooth, subhyaline.
Habitat: HOSTS: Freycinetia baueriana subsp. banksii, unidentified wood.
MATRIX: Corticated twigs, roots.
Notes: NOTES: The New Zealand collections were compared with the Sri Lankan specimens of R.
arcuata collected by Petch. The subiculum and the Dematophora anamorph as well as the
ascospore shape and the short germ slit are identical. In addition, the statistical analyses
revealed that there are no differences among the collections regarding quantifiable characters
such as size of stromata, ascospores, or the ascus apical rings.
Macroscopically R. arcuata resembles R. necatrix Prill., but its ascospores are statistically
significantly larger (Fig. 9A). Roger (1953) and Saccas (1956) reported R. arcuata among
other host plants on roots of Camellia sinensis (L.) Kuntze and Coffea arabica L., damaging
them severely. The stromata and ascospore sizes are similar to those reported for the New
Zealand and the Petch specimens, but both authors describe a germ slit about twice as long.
According to Saccas (1956) the geographical distribution of R. arcuata is confined to tropical
and humid regions of Africa and Asia. In fact, the Petch specimens are from a mountainous
region of Sri Lanka. In New Zealand, R. arcuata was mainly collected in the northern part of
the North Island in national parks and state forests with subtropical vegetation and has not
been reported from cultivated trees.
There are some specimens in PDD labelled as R. necatrix, with only the Dematophora
anamorph present. They originate from Narcissus sp., Vitis vinifera L., Juglans
regia, Malus
sylvestris L., and Malus x domestica Borkh. (PDD 50725, PDD 49923, PDD 26403, PDD
17391, PDD 24985, respectively) and may be, indeed, R. necatrix, as these are typical hosts
for this species. In fact, root diseases caused by R. necatrix were reported by Bösewinkel
(1977) and are listed in the New Zealand Plant Disease Database. Most Rosellinia species
reported as severe root pathogens have ascospores similar to those of R. arcuata or R.
necatrix, slender with tapering ends, short germ slit, and slimy sheath, varying only in size.
Molecular biological studies are needed to confirm or reject conspecificity for suchtaxa.
Rosellinia asperata Massee, R. bothrina Berk. & Broome, R. desmazieresii (Berk. & Broome)
Sacc. var. acutispora Theiss., R. gigantea Ellis & Ever., R. pepo Pat., and R.
puiggiari Pat.
possess ascospores with a morphology similar to those of R. arcuata.
Examination of the type materials from the herbaria FH, K, NY, and PACA revealed that
character combinations such as stroma shape and size and ascospore size do not match those
of R. arcuata (L. E. Petrini unpubl. data).
|