|
 Cryptohymenium pycnidiophorum Cryptohymenium pycnidiophorum
BiostatusPresent in region - Indigenous. Endemic
Images (click to enlarge)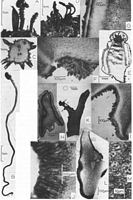
Caption: Fig. 1: A-C. Cordierites acanthophora: A. Apothecia as found in nature. - B. Apothecia on long stipe. -
C. Transverse section through stipe showing
peripherally situated pycnidia. (PDD 46956). -
D-G. Cryptohymenium pycnidiophor | 
Caption: Fig. 3: A-G. Cryptohymenium pycndiophorum: A. Diagram of nine apothecia. - B Section showing "epithecium", tips of asci and paraphyses (Melzer's reagent).
- C. Asci, ascospores and paraphyses (Melzer's reagent). - D. Median longitudinal section |
Article: Samuels, G.J.; Kohn, L.M. (1987) [1986]. Ascomycetes of New Zealand. 7. Some bizarre, inoperculate discomycetes. Sydowia 39: 202-216.
Description: Apothecia up to 10 cm long, arising from decaying herbaceous debris and soil, gregarious,
consisting of a long, slender, scaly stipe 1-1.5 mm diam x up to 15 cm
long and a conical, 2-2.5 mm long cap. Cap separated from stipe by a distinct groove; consisting
of a hymenium surmounted by a globose, multiloculate
pycnidium delimited from hymenium by a groove; hymenium covered by a cellular epithecium that
eventually flakes off; stipe growing through substrate as a
rhizomorph. Stipe unbranched or branched one to several times near the apex to produce a
panicle of stipitate caps; coarse, dark brown to black, unbranched
hyphae arising from base of stipe, growing through substrate. Entire cap at first covered
by an epithecial tissue ca. 100 µm thick and consisting of an outer region
30-50 µm wide composed of brick-like cells 10-20 µm long x ca. 10 µm wide oriented with long axis
perpendicular to asci, walls ca. 1.5 µm thick, brown; cells
of inner 20-30 µm circular to square in outline, 10-15 µm across with walls ca. 1 µm thick,
lightly pigmented; cells of inner layer breaking down; epithecial tissue
eventually flaking away to reveal the hymenium; cells of epithecium continuous over pycnidium
and with cortex of stipe.
Asci (130-)150-196(-220) x 9-12(-14) µm, narrowly clavate, tapering to truncate base,
pore J + Melzer's, cylindrical, ca. 2 pun long x 3 µm wide; ascospores
generally forming in the upper (25-)90-140(-150) µm of the ascus. - Ascospores
(21-)23-28(-40) x (3-)4.2-5.4(-6) µm, partially biseriate; fusoid to subfusoid,
containing 2-3 irregularly shaped guttules, smooth, hyaline. Paraphyses equal to or
slightly longer than asci consisting of branching, septate, 2-3 µm wide stalk
and subglobose to clavate tip; tip 10-20 µm long x 8-10 µm wide; tips of paraphyses
encrusted in amorphous, brown material; encrusting material turning red in
3% KOH. Subhymenium 60-70 µm thick, consisting of compact, interwoven hyphal cells with
brown walls; irregularly shaped accretions of brown pigment
scattered throughout the subhymenium. Medullary excipulum well developed, consisting of
brick-like cells 20-40 µm long x 7-10 µm wide, long axis oriented
vertically, walls ca. 1 µm thick, lightly pigmented. Ecta1 excipulum not well developed,
30-100 µm wide, consisting of square to rectangular cells 15-25 µm long
x 7-15 µm wide, walls dark brown, ca. 1.5 µm thick, long axis of cells parallel to long
axis of asci, continuous above with epithecial tissue and below with cortex
of stipe. Tip of cap separated from body of cap by a constriction, cells of tip textura
angularis, 5-10 µm in greatest dimension; labyrinthiform, conidiogenous
locules forming within tissue of the tip, lined with conidiogenous cells. Conidiogenous
cells arising directly from surrounding cells, ampulliform, tapering from
base to tip, 10-15 um long x 3-4 µm wide at base x 1-1.5 µm wide at tip, phialidic,
monoblastic, tip thickened, collarette not flared, light brown. - Conidia 2-3 x
2-2.5 µm, globose to subglobose, lacking a basal abscission scar, unicellular, hyaline.
Stipe composed of cortex and medulla; cortex 50-70 µm wide, composed
of 4-5 layers of square to rectangular cells 10-20 µm long x 7-10 µm wide, walls ca.
1.5 µm thick, dark brown, with long axis parallel to long axis of stipe,
aggregates of cells coming off as scales. Medulla composed of an inner and an outer
region; outer medulla adjacent to cortex, ca. 150 µm wide, cells rectangular,
30-45 µm long x 8-11 µm wide, walls very pale brown, with long axis parallel to long
axis of stipe; merging at exterior with cortex and at interior with inner
medulla. Inner medulla ca. 300 µm wide, composed of hyphal elements with cells 40-100 µm
long x 4.5-6.0 µm wide, walls ca. 1.5 µm thick, pale brown, with
long axis parallel to long axis of stipe.
Notes: Cryptohymenium pycnidiophorum is certainly among the most bizarre of the discomycetes.
Its fructification appears to be a rhizomorph the tip of which
expands to form, simultaneously, the anamorph and the teleomorph. The hymenium is surmounted by the
pycnidium and is covered with a layer of
pseudoparenchymatous cells that is continuous below over the rhizomorph. The hymenium is exposed
when this layer of cells flakes off.
Although the fructification of C. pycnidiophorum is unusual, the dark
pigmentation of the receptacular tissues, the green hymenium and the angular cells of the
ectal excipulum, and the eustromatic pycnidium combine to indicate that the genus is
a member of the Helotiales family Dermateaceae (KORF, 1973). This
species bears some similarity to Atropellis Karsten (REID & FUNK, 1966).
The hymenium of species of both genera are covered with a tissue that is continuous
with the ectal excipulum and that flakes away at maturity. In Atropellis the
tissue is hyphal whereas in Cryptohymenium it is distinctly cellular. The covering of
Atropellis dehisces through a preformed opening but such an opening was not
seen in the covering of C. pycnidiophorum. Tips of the paraphyses in both genera
are ensheathed in amorphous material. This material is green in Cryptohymenium,
it turns red in 3% KOH and a red pigment is released in that reagent. The
material of Atropellis is usually blue-black and a blue-green pigment is
released in 3% KOH. The pycnidium of Cryptohymenium is eustromatic (ss. Sutton,
1980) and is anatomically similar to pycnidia of Fuckelia BON., some
species of which have been linked to Atropellis (KORF, 1973). We do not know whether
the conidia of the Cryptohymenium anamorph are disseminative or spermatial.
Despite diligent efforts, we have not been able to establish a connection to a
definite substrate. We note the near constant association of the species with living
roots of the usually mycorrhizal genus Nothofagus. It is tempting to suggest
a mycorrhizal role for C. pycnidiophorum.
|