|
 Clavulina urnigerobasidiata Clavulina urnigerobasidiata
BiostatusPresent in region - Indigenous. Endemic
Images (click to enlarge)
Owner: J.A. Cooper | 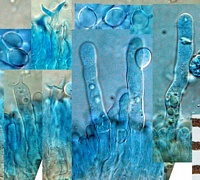
Owner: J.A. Cooper |
Article: Petersen, R.H. (1988). The clavarioid fungi of New Zealand. New Zealand Department of Scientific and Industrial Research, Bulletin 236: 170 pp. Wellington:.
Description: Fruit bodies up to 45 x 20 mm, branched, obpyramidal to fusiform in general outline, gregarious to cespitose, arising from a mycelial mass involving soil substrate but very little visible mat. Stipe discrete, up to 20 mm long, tan ("tawny olive"), or fruit body branching from the base, often below substrate level so as to appear cespitose, flattened somewhat to lobed in cross section. Branches in 1-3 ranks, flattened, dull fleshy tan to tan ("cinnamon buff', "tawny olive") when young, dull violaceous tan ("wood brown") when mature; internodes diminishing gradually; axils narrowly rounded to acute. Apices subcristate to narrowly lobed or minutely mitten-shaped when mature, cristate only when young, paler than branches ("avellaneous").
Macrochemical reaction: FCL = darkening but not green.
Tramal hyphae of stipe and upper branches 2.5-7 gm diam., hyaline, clamped, parallel. Subhymenium
scanty. Hymenium not significantly thickening; basidia (Fig. 62) 36-41 x 7.5-8.3 gm, clamped, subcylindrical to subtly suburniform (see commentary), golden tan under bright field, yellow¬refringent under phase contrast; contents multiguttulate with one large guttule predominating; sterigmata 2, slender, up to 6.5 gm long, divergent¬curved; post-partal septation absent. Cystidia (Fig. 63) unobtrusive, 30-40 x 6.8-9 l m, elongate barrel-shaped to subcylindrical, to lobed or branched, not septate, hardly protruding from hymenium, clamped.
Spores (Fig. 64) 7.9-9.7 x 6.8-8.6 gm (E =1.00-1.17; E, = 1.10; L'° = 8.57 gym), globose to subglobose, thin¬walled, uniguttulate, very pale beige under bright field; hilar appendix papillate.
Notes: The tan colour is not restricted to hymenial elements, although it is most pronounced there. The stipe and lower branches are also tan, but I cannot discern whether the colour is in the cell contents or cell walls. If one concludes that the walls are pigmented, then subg. Fusco-clavulina Corner must be considered. None of the taxa therein fit this collection, however. Likewise, I cannot find a taxon in subg. Clavulina which matches.
The flesh is white, contrasting with the surface. This may be what Corner (1970) reported as Clavulina mussooriensis from New Zealand. Specimens under that name from India in TENN show cystidia emergent and often septate, basidia with post-partal septation, and subglobose to ellipsoid spores (El =1.28 gm). I have not seen the New Zealand material at CGE, so I cannot judge its identity.
Basidial ontogeny is as follows: very young basidia are bullet-shaped. The first basidium of a cyme forms a clamped septum at this early stage, but subsequent basidia, which arise as branches just below the subbasidial swelling of the previous basidium, do not form such a septum until later in their development. The basidial initial elongates somewhat, to a length of 17-22 gm and a rather constant diameter. When this length range is attained, elongation apparently slows or stops, and two processes occur: (i) the initial inflates somewhat to become elongate barrel-shaped rather than digitate; and (ii) refringent yellowish guttules begin to appear in the basidial lumen. From relative numbers of such young basidia versus mature basidia, I conclude that this stage is rather long, i.e., that further maturation is postponed. When elongation is reinitiated, the apical portion of the maturing basidium is somewhat narrower than the initial from which it proceeded, and although some circumferential enlargement occurs apically, most mature basidia show a median constriction resulting from this two-stage maturation. For a time, the oil droplets remain in the basidial base, but eventually migrate distally, increasing in number and coalescing into larger individuals. Mature basidia often show one very large apical guttule, as well as many smaller individuals.
Sterigmata are relatively short and slender for the genus, but otherwise normal. After spore discharge, post-partal septation seems absent, but effete basidia are persistent. This mode of basidial maturation, so far as I know, has not been reported in this genus. Corner (1950,1970) understandably was concerned with other more strikingly anomalous basidial characters, such as sterigmata number and post-partal septation. Some of his figures (Corner 1950, e.g., text figs 25,133) suggest that the bullet-shaped initial expands as described above.
Conversely, this ontogenetic progression is typical of Multiclavula (Petersen 1967a) in the clavarioid fungi. It is best described as "suburniform" in the sense of Oberwinkler (1982). This basidial ontogeny is much better known in the corticioid Homobasidiomycetes. Boidin (1958) merely indicated that Sistotrema Fr. (= Trechispora seas. str. Boidin) produced urnigera-type basidia, and showed stichic basidial nuclear spindles in meiosis. This character coupling was mentioned by Donk (1964b), but not emphasised, and in a list of genera with stichic basidia (Donk 1964b; p.222) no members of the corticioid group was included. Hubbard & Petersen (1979) reported the coupling in Multiclavula (see below under Multiclavula Petersen). Using Oberwinkler's (1982) hypothesis, the urnigera basidia of Sistotrema can be compared favourably with those of Hyphodontia Eriksson. Eriksson & Ryvarden (1976) compared Hyphodontia with Hyphoderma Wallr., and under the latter name (Eriksson & Ryvarden 1975), several rather striking similarities to Clavulina may be found. Cystidia are very similar, includingboth septate (and clamped) and non-septate forms, constricted, "suburniform" basidia, and spores with copious oil inclusions. Conversely, I can find no literature on basidial nuclear behaviour in Hyphoderma or related genera (but I have hardly searched), except some illustrations by jiilich (1974) of H. setigerum (Fr.) Donk which show a 4¬nucleate basidium, the nuclei of which are low in the basidium. These may be residual nuclei, however, and therefore not indicative.
Two younger basidia are depicted, again in 4-nucleate stages, and again the nuclei are low in the basidium, a location rather indicative of stichic behaviour (cf., Boidin 1958, Donk 1964b). Finally, the circumscription of Hyphoderma offered by Jiilich & Stalpers (1980) is not at odds with the microscopic characters of Clavulina, although the diversity of all structures is much wider in the former than in the latter.
The above are mentioned because of the traditional taxonomic isolation of Clavulina. With the possible exception of the resupinate genus Clavulicium Pilat which has been likened to Clavulina by some authors (Parmasto 1968), Clavulina has stood alone since its segregation from other clavarioid genera by Schroeter (1889). Donk (1964b) placed it in a monogeneric family, the Clavulinaceae, compared to Canlharellus L.: Fr. and Hydnum L.: Fr. but far removed from them. The most pressing need is for observation of basidial nuclear behaviour in the Hyphodermoideae. Cystidia in this taxon can be distinguished from young basidia only because young basidia include discrete oil guttules, while the entire contents of similar cystidia are refringent. Longer cystidia present less trouble, even though they rarely protrude from the hymenium.
|