|
 Limacella whereoparaonea Limacella whereoparaonea
BiostatusPresent in region - Indigenous. Endemic
Images (click to enlarge)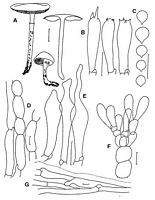
Caption: Limacella wheroparaonea: A, habit and section of basidiocarp (after Stevenson's drawing);
B, basidia; C, spores; D, hyphae from annulus margin; E, pleurocystidia; F,
elements from the hymenium and subhymenium; G, gelatinous pileipellis. Sca |
Article: Ridley, G.S. (1993). Limacella macrospora Stevenson and L. wheroparaonea, a new species, from New Zealand (Fungi, Agaricales, Amanitaceae). Australian Systematic Botany 6: 155-159.
Description: Pileus 18-30 mm diam., convex to plano-convex
with even margin, not hygrophanous, 'red-brown' or dull brick, moist or viscid
at first, drying sparsely pubescent, margin exceeding lamellae and non-sulcate.
Lamellae crowded, sinuate, subventricose, 'creamy' or very pale buff, margin
entire. Stipe 25-50 x 3-5 mm, cylindrical, solid, rather fragile, pale luteous
buff and flecked with umber below annulus (this latter feature is portrayed
in the water colour and possibly represents litter particles), above annulus
sericeous, texture below annulus not recorded, annulus arachnoid and fugacious.
Context very pale buff.
Spore print not obtained. Basidiospores (12/1),
(3.5-)4-5(-6) x (3.5-)4-5(-6) µm, Qm 1.01, Q 1.00-1.11, globose,
hyaline, neither amyloid, dextrinoid nor metachromatic. Basidia 22-25 x 5-6
µm, with clamp connections, 4-spored, rarely 1-2 spored, then basidiospores
generally 6 µm diam. Lamellae edges fertile, cheilocystidia absent. Pleurocystidia
26-39 x 5-7.5 µm, rostrate, with rostrum straight to flexuose or rarely unequally
bifurcate, thin-walled, hyaline, with clamp connections, standing 10-15 µm above
the general level of hymenium. Hymenophoral trama divergent, hyphae 12-15 µm
diam., with sparse oleiferous hyphae, 2-3 µm diam. Subhymenium of small inflated
cells and sub inflated short hyphal segments, to 15 µm diam., hymenial elements
arising from either type of cell, arranged in a branching structure, roughly
perpendicular to the hymenial surface, with elements becoming larger towards
the central stratum. Pileipellis a transition between an ixocutis and an intricate
ixotrichoderm, made up of repent to ascending hyphae with non-specialised terminal
cells, 5-7 µm diam., with clamp connections: pigment pale umber, plasmatic,
leaching rapidly in ammonia solution. Subpellis and pileitrama regular to irregular,
with sparse oleiferous hyphae 2-3 µm diam., acrophysalides absent. Stipitipellis
a cutis made up of interwoven hyphae, 6-9 µm diam., with clamp connections,
with non-specialised terminal cells. Stipititrama regular, hyphae macroscopic
1 cm, microscopic 10 µm. to 10 µm diam., sparse oleiferous hyphae to 8 µm diam.,
with clamp connections, acrophysalides absent. Annulus made up of parallel hyphae,
3-10 µm diam., which increase in diam. towards the annulus margin, near margin
septa more numerous and hyphae inflated to form chains of cylindrical, subglobose
cells, 10-32 x 8-15 µm, eventually disarticulating.
Habitat: Under 'broadleaved scrub' (from specimen
label). Known only from the type locality, southern North Island, New Zealand.
Notes: Etymology and Pronunciation
Whero, red + paraone, brown (whero'para'one),
from Maori, referring to the colour of the pileus. The phonetic nomenclature
follows the conventions outlined in Sykes (1982). The initial wh represents
a voiceless consonant, in Maori, equivalent to w, and is pronounced by emitting
the breath sharply between the lips (Williams 1975).
Notes
In section the overall appearance of the
hymenophoral trama is distinctly divergent and of the Amanita type. Bas
(1969) defined this type as composed of acrophysalides, produced on short lateral
hyphae, and feeding hyphae. The hymenium is produced from the feeding hyphae.
The expansion of the gills as the basidiocarp
grows is achieved by the inflation of the acrophysalides. However he notes that,
in some members of Amanita section Lepidella, the acrophysalides are
scant or lacking and that in these species expansion of the gill is achieved
through the inflation of the feeding hyphae. This feature he considered to be
primitive. Microscopic examination of the hymenophoral trama of L. wheroparaonea
failed to reveal any acrophysalides. Instead, it is constructed of inflated
feeding hyphae, as described for Amanita vittadinii (Mor.) Vitt. and
A. lilloi Sing. (Bas 1969). It is interesting to note that Bas described
the same inflated feeding hyphae in the stipititrama of these same species of
Lepidella and in Limacella guttata (Fr.) Konrad & Maubl.,
but no such inflated hyphae could be found in the stipititrama of l. wheroparaonea.
Thus, if Bas's contention is correct that the lack of acrophysalides is primitive
and that the section Lepidella and Limacella are the most primitive
of the Amanitaceae, then the lack of inflated elements in the stipititrama of
l. wheroparaonea marks it as one of the most primitive species in the
genus.
Bas (1969) lists 13 trends in the evolution
of Amanita of which a number, those not concerned with the structure of the
volva, are applicable to Limacella and support the primitiveness of L.
wheroparaonea in the Amanitaceae. These primitive features are the poorly
differentiated inflated trama elements, abundant clamp connections, non-sulcate
pileus margin, thin-walled, globose spores and probable saprophytic, as opposed
to mycorrhizal, lifestyle.
Singer (1986) divided Limacella into
two sections, section Lubricae and section Limacella (= Viscidae
(Smith 1945)), the former with a viscid or glutinous stipe and the latter with
a dry stipe. L. wheroparaonea belongs to section Limacella as
it has a dry stipe. Using Smith's key l. wheroparaonea is best placed
near l. glioderma (Fr.) Earle and L. delicata (Fr.) Earle, sharing
similar red-brown coloration and globose spores in the 3-5 µm range. Limacella
wheroparaonea is unique, as it appears to be the only species in the genus
to have pleurocystidia. However, as our knowledge of this genus is primarily
based on the North America flora, the presence of pleurocystidia may prove to
be more widespread as other flora are investigated.
The collection data give little information about this species habitat;
however, Viggers (1978) describes the vegetation of Jubilee Park as forest,
cut over in the late 19th century, and still containing Alectryon excelsus
Gaertn., Metrosideros robusta A. Cunn., Beilschmiedia tawa (A.
Cunn.) Kirk, Dacrycarpus dacrydioides (A. Rich) De Laubenfels and Laurelia
novae-zelandiae A. Cunn. Thus it is probable that the distribution of L.
wheroparaonea will be linked with the distribution of podocarp-broadleaf
forests. In comparison, all of the endemic species of Amanita are found
in mycorrhizal association with Nothofagus Blume, Leptospermum scoparium
Forster & Forster f. and Kunzea ericoides (A. Rich) J. Thompson (Ridley
1991).
|